
Across my first three posts on autopoiesis I have contrasted strict and looser definitions of the concept. The stricter definitions are more logically consistent; they hold together rigorously. The looser definitions apply to increased real and imaginary cases; they have greater extension.
It would be a mistake, though, to think that the opposition of strict and loose is between cohesion and applicability. These criteria can distinguish them, but there is something deeper and more important at work.
The distinction between strict and loose concepts of autopoiesis is about explanation. For the strictest definitions the role of the concept is, above all, to explain how some processes can sustain autonomy, self-production, change and unity. For the looser definitions the role is, above all, to explain a range of phenomena as autopoietic.
For the stricter definitions, the prime focus is internal to autopoiesis. It has the guiding question: ‘How can autopoiesis be explained as a lasting and consistent process?’ The looser definition is determined more by the questions: ‘Which of these phenomena are autopoietic? and ‘What does this explain about them?’ The main concern is external.
I have insisted on the relative nature of these roles; ‘above all’ one way or another. This is because they blend. To a lesser extent than the more empirical approaches, specific phenomena are still included in the stricter ones. Furthermore, the strict definitions include the claim that autopoiesis is equivalent to life, thereby including all living things under the concept, in principle.
All the looser approaches are informed by necessary and logically consistent features of autopoiesis, though these don’t dominate explanations. Where a particular empirical case calls for refinement of the concept, these approaches bend the pure definition in order to extend its reach.
This relativity does not mean that the differences between modes of explanation are insignificant. On the contrary, different approaches to explanation define contrasted ways of thinking about autopoiesis.
The distinguishing feature of Maturana and Varela’s work isn’t that it came first, but rather that it is strict and seeks – first and foremost – to explain an inner consistency and autonomy in a logical and rationalist manner. Whereas Evan Thompson’s study of autopoiesis stands out because of its empirical and phenomenological inquiry around autopoietic phenomena.
Thompson’s work will be the main topic of this post. I have taken his studies as the main example of the looser approach. The relativity of the two poles of strict and loose can be seen not only in his frequent references to Maturana and Varela’s definitions and arguments, but also in Thompson’s analysis of their approach to phenomena; particularly to cells, cognition and life.
The point here is neither to judge different definitions of autopoiesis as better or worse, nor to attack either definition as weak or flawed. It is to chart their different consequences for a semiology of autopoiesis and for a critique in terms of timed logic: how autopoiesis works as a sign and, eventually, its relation to ideas of time.
These contrasts were already a factor of my previous post on Claire Colebrook’s arguments on time and autopoiesis. Her points on signs and images applied more strongly to the looser definition of autopoiesis and did not have much traction against the stricter version.
The function and necessity of a boundary for an autopoietic process is a good way of understanding the difference between modes of explanation. To explain the existence of a category of empirical cases of autopoiesis, looser approaches view physical boundaries as necessary for the identification of a phenomenon as autopoietic. This is not the case for strict definitions, where observable physical boundaries are not primary conditions for autopoiesis, but rather consequences of it for some cases.
The difference is between an abstract concept of self-protection that follows from unity and autonomy, and only involves a material boundary under certain conditions, and an empirical observation of boundaries as condition for the identification of autopoiesis and explanation of its sustainability.
In physical (biological and chemical) environments an autopoietic process seems to maintain autonomy and identity only if it has a boundary of the kind we might recognise as protective and containing, such as a wall, skin, membrane or sack. These must be sufficiently permeable to allow for energy exchanges, yet they must also defend inner processes from outer threats and control.
According to pure definitions around unity, a protective process is a consequence of the distinction between inclusion into and exclusion from unity. It also follows from the separation between an autonomous process and threats to it. Finally, given the requirement for external energy and the challenges it presents for autonomy, the protection is defined by permeability restricted to necessary energy sources and other external requirements such as information. Such a discriminating limit must remain inviolable by anything that might undo autonomy.
The differences between the empirically derived and pure definitions are therefore between contained identity and protected unity as ways of maintaining autonomous self-production. Less precisely, this can be summed up as the difference between an observable boundary round an identified process or set of processes (a cell) and a unified network of protected processes (an evolving electrical grid, or a cell thought of strictly as a network of processes).
These differences play out in terms of relative and absolute autonomy. For the looser definitions, an autopoietic system can be controlled from the outside so long as its core independent processes are relatively unaffected. A living thing can be breached and affected from the outside, yet remains autopoietic so long as this breach is contained. For the strict definitions, if there is such control, then autopoiesis ceases.
This distinction also turns on the contrast between identity and unity. In the loose version, these terms are used together to determine unified self-identity, including a wide range of empirical cases and their variations over time and individuals. In the strict version, unity is all-important and defined as an absolute term, in the sense that there is either unity or not, independent of any further mitigating conditions.
For the loose version, an autopoietic process is malleable and variable, depending on the preservation of some kind of self-produced identity for a life form, like the growth of a named living thing – a self – through a series of phases.
In the following passage Thompson brings together ideas of autopoietic network, individuality, identity, system, boundary and self. The result is a rich and complex image of autopoiesis, allowing for its extension from cells, to individuals, minds and selves. This is at the expense of tensions and contradictions between these terms, when compared to more strict definitions; for instance, around networks and boundaries, identity and unity, and autopoietic processes and selfhood:
An autopoietic system is a circular network whose constituent molecular processes both produce and embody that network. Individuality in this case corresponds to a formal-self-identity – to an invariant dynamic pattern that is produced, maintained, and realised by the system itself, while the system undergoes incessant material transformation and regulates its external boundary conditions accordingly. An autopoietic system is thus an individual in the sense that begins to be worthy of the term self.
Evan Thompson (2007) Mind in life: Biology, Phenomenology, and the Sciences of the Mind Cambridge MA: Belknap Press of Harvard University Press, p 75
Against this intermingling of concepts, the strict version fixes autopoiesis to the preservation of a tightly defined process of autonomous production. There is no plasticity or variation at the abstract level defining that process, in contrast to the potentially very high degree of change in its products.
According to the strict version, unity is defined by an abstract plan. It cannot have the completeness and richness identified with a self, because this always comes at the price of an end to absolute autonomy, due to the ways a self is created in conjunction with others; that is, by giving degrees of control over to them.
If we take autonomy in a strict sense, a functioning ethical society of autopoietic selves is a nonsense, because such a society must rest on trusting others over oneself. I willingly give you control over my development.
This does not mean that the strict definition can’t be consistent with societies or selves. It depends on how the autopoietic process is kept at a distance from social contracts and mutual responsibilities, either written or implied, and on how the idea of self is restricted to avoid claims on the self by others, such as Sartre’s ‘look’ and the hold of the other’s gaze over the self:
The Other is present to me everywhere as the one through whom I become an object.
Jean-Paul Sartre (1992) Being and Nothingness, New York: First Washington Square Press, p 373
If autopoiesis takes place in societies it cannot be at the level of an autopoietic self, understood in the full sense of individual human, but rather through the coexistence of autopoietic abstract plans that come into and out of existence within a shared society, when social requirements give space for, or remove, absolute autonomy. Your autonomous power to create yourself has been cancelled.
Here, ‘to give space’ cannot mean ‘to continue to allow’, since any monitoring would imply a minimal external control and potential for intervention. For instance, a practice would not be autonomous if it were subject to self-censorship induced by an external threat.
A self can maintain its identity through varied phases, some fast and some slow, some smooth and some staccato, like an individual animal life with its many social and environmental interactions over seasons and life stages. A strict autopoietic process associated with its defining unity is only on or off. It implies jumps between different autopoietic processes, moments of preservation and extinction, and lines of independent abstract plans described according to their passage from one to another.
The emphasis on observability in these distinctions refers back to my opening comments on the looser definitions of autopoiesis as explanations of empirical phenomena. In such cases, the boundary is observed as containing an identified autopoietic process. This can take place on many different scales from microscopic to macroscopic. There are also difficult cases such as insect colonies, whose status as autopoietic is unclear:
Autopoiesis is the paradigm case of biological autonomy for two reasons. It is empirically the best case, and it provides the core “biologic” of all life on Earth. To qualify as autonomous, however, a system does not have to be autopoietic in the strict sense (a self-producing bounded molecular system). An autopoietic system dynamically produces its own material boundary or membrane, but a system can be autonomous without having this sort of material boundary. The members of an insect colony, for example, form an autonomous social network, but the boundary is social and territorial, not material.
Evan Thompson (2007) Mind in life: Biology, Phenomenology, and the Sciences of the Mind Cambridge MA: Belknap Press of Harvard University Press, p 44
Thompson’s discussion leans heavily on boundaries as necessary for autopoiesis. Even where he considers examples with no material boundary, there is an insistence on social and territorial boundaries, both of which remain observable as features of territories and societies. When he uses the notion of ‘strict’ autopoiesis, it is not in the pure logical and rationalist sense, but rather as a description of empirical biological processes as a paradigm that other living organisations tend towards:
There is a basic organisation of life, and its paradigm and minimal case is to be found in the single cell. A single-cell organism is a self-making or self-producing being. Self-production is different from reproduction: In reproduction, a cell divides in two; in self-production, a cell continuously produces itself as a spatially bounded system, distinct from its medium or milieu. What is remarkable about self-production is that every molecular reaction in the system is generated by the very same system that those molecular reactions produce.
Evan Thompson (2007) Mind in life: Biology, Phenomenology, and the Sciences of the Mind Cambridge MA: Belknap Press of Harvard University Press, p 92
Thompson uses ‘paradigm’ frequently and at key points in Mind in Life. The word is tricky, because it can mean ‘typical example of’ (Paris is the paradigm of capitals) or ‘model for’ (single span wings became the paradigm for airliner design). The difference is significant because a model has a strong claim over what it is a model for (always start the design with a single wing span), whereas a typical example does not (Astana does not exhibit power in the same way as Paris).
Close examination of the arguments where Thompson uses the term paradigm shows that he is using it in the sense of model, with dominant features and model cases: ‘the paradigm for interaction with a heteronomous system is input/processing/output’ (p 38); ‘the paradigm for interaction with an autonomous system is a conversation’ (ibid); paradigm as ‘best case’ and ‘biologic’ (p 44); ‘paradigm case of dynamic co-emergence’ (p 65); ‘paradigm and minimal case’. This again illustrates the empirical and biological sources of his definition of autopoiesis. It follows, above all, from an observed model taken from the biological sciences, then combined with other logical and rational theories of autopoiesis.
The empirically derived definition can be included under the strict logical one, if the protection process from the latter is by observable containment and if unity is identifiable over time. However, the reverse does not hold, since unity is not always identity (in a unity of fully transformational processes, for instance) and since protection need not take the form of an observable containing boundary (for a widely distributed and changing network, for example).
A spy ring is a helpful analogy for this latter kind of self-generated protection. Spies might be spread throughout countries such that there is no observable material, physical or social boundary to the ring. Spying often takes advantage of the unreliability and ambiguity of social boundaries, as in the establishment credentials used as cover by the Cambridge Spy Ring. A double-agent operates between (at least) two social groups, observably belonging to both, depending on the observer’s perspective.
In order to function, a spy ring should be autonomous, in the sense of independent of foreign influence and control. It should also be permeable to useful information, in one direction, and disinformation and rewards for contacts in the other.
The abstract protective process of successful spying is constituted by rules, practices and technologies, such as codes, secret networks and disguise. It is not observable in usual ways seeking to detect a continuous separation between inside and outside. The criterion of an uninterrupted and observable boundary will fail for any distributed system of autopoiesis of this kind. A spy ring is not broken by piercing its boundary, but by understanding its methods of communication, concealment, recruitment, information-gathering and funding.
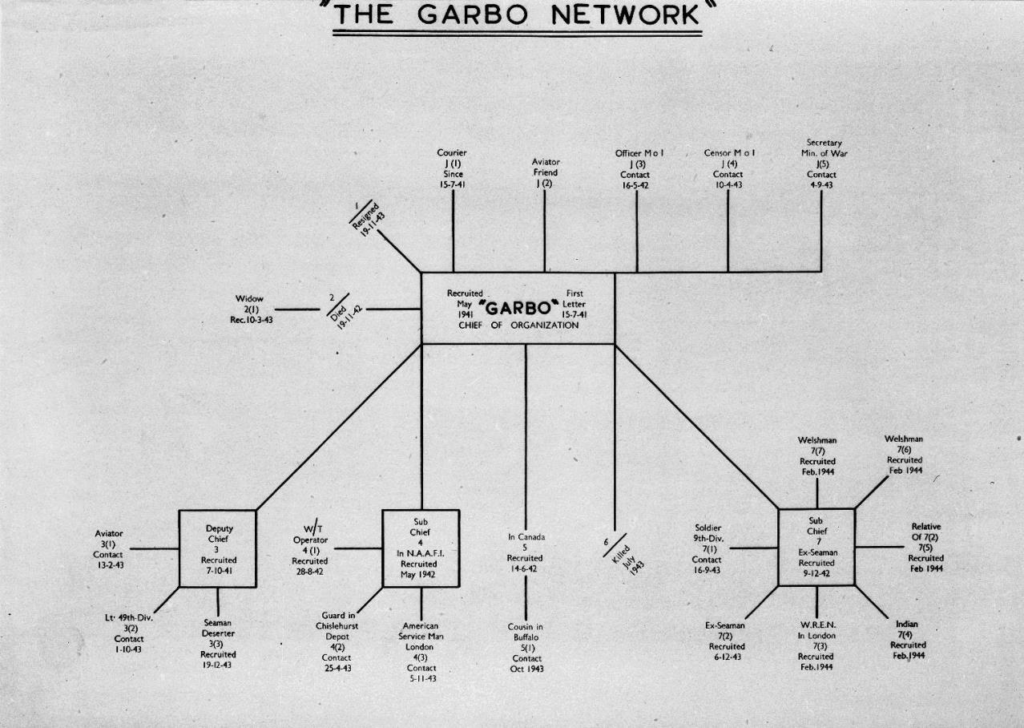
A distributed method of protection is still discernible, if we change from a search for visible boundaries to inquiries into closed circuits between separate components; for instance, in the study of the information flows linking spies. In this shift from containment to inviolability, the concept of an autopoietic boundary shifts from empirically observed enclosures to semi-closed networks.
Analogies can be useful tools for explanation and understanding, but they have the potential to foster illusory command. The analogical claim to parallels between different subjects is abstract and high level. It concerns similarities between structures, rather than shared detailed processes.
These similarities can help us understand how things function. For instance, like novelists, academic researchers keep extensive notes and work from them. Notes are often important. Yet there is always the danger of distorting and misrepresenting the underlying processes. Beyond note-taking, academic researchers are far from novelists.
Despite this risk, I will pursue the analogy between abstract boundaries and spy networks to suggest some differences in threats between containing boundaries and semi-closed networks. Furthermore, over the course of my semiology of autopoiesis, I want to study the relation between pure concepts, signs, metaphors, analogies, models and examples. Semiology is well-placed to follow the effects of slippage between these techniques, because signs operate in all of them, with connections and implications that can be unpacked thanks to a theory of signs.
A continuous physical boundary is most vulnerable to a single type of threat, a breach. Furthermore, the biggest risk is at an allowed crossing point. The weakest point is where a necessity has to cross the boundary, hence the concern in autopoiesis about corrupted energy sources such as poisons. In contrast, for a network, there are potentially different kinds of threat for each node of the network and for each interchange between two or more nodes.
In a spy ring, every agent is for turning and in many different ways (financial pressures, sexual desires, ideological beliefs and disillusion, jealousies and fears, psychological flaws and strengths, ambition and fatigue, academic connections and settings, and family ties). Each communication is interceptable, decodable and fakeable. Weak points are everywhere. In the Garbo ring pictured above, the perceived weak point was familial and psychological, leading to a deeply unfair and potentially counter-productive defensive measure.
For boundaries, the guiding defensive idea is to build the best possible wall, skin, sack, envelope or membrane. For protective networks, the more demanding idea is that any network is only as strong as its weakest point. It is worth following the link in the spy network picture to learn from Richard Dunley’s sensitive historical studies of the kind of breaches that might occur, for instance, if a code or technique is broken.
When autopoiesis and its cases and paradigms serve as a sign, as they do when the concept becomes a model or value to follow, there is a great deal at stake about whether it must involve an observable boundary and, more significantly, whether any process should be thought of as satisfactorily bounded. Firstly, there can be errors of misattribution; about weak points, for instance. Secondly, these errors can become strategic; when we underestimate the number and types of threat to a process.
It matters whether we focus on boundaries or more diffuse and distributed protective processes, because these also affect what we take to be a threat and how it is dangerous. It could be that the concept of boundary is exactly the wrong image to foster and live by, even for autopoiesis.
For the abstract and more strict definition of autopoiesis, no empirical observation can lay claim to have found a general and necessary form for autopoietic boundaries. It has merely detected a manner in which the limit between unity and its outside is manifested. In theory, this limit can take any form so long as it satisfies conditions set by autonomy, unity and exchanges across the limits preserving them.
The following passage from Mind in Life gives an empirical account of the necessity of a cell’s boundary. As a description tied to a particular science, to its probability, acceptance, scope, epoch, counter-theories and experiments, it cannot be used to justify the necessity of boundaries for autopoiesis:
A cell stands out of the molecular soup by creating the boundaries that set it apart from what it is not and that actively regulate its interactions with the environment. Metabolic processes within the cell construct these boundaries, but the metabolic processes themselves are made possible by those very boundaries. In this way, the cell emerges as a figure out of a chemical background. Should this process of self-production be interrupted, the cellular components no longer form a unity, gradually diffusing back into the molecular soup.
Evan Thompson (2007) Mind in life: Biology, Phenomenology, and the Sciences of the Mind Cambridge MA: Belknap Press of Harvard University Press, p 46
The metaphor of ‘standing out from’ captures the independent identity of the cell and not its unity. It isn’t necessary to have boundaries to be distinguished from a ‘soup’ or to be a distinct figure against a ground. To stand out thanks to a boundary is a particularly stark form of independent identity, but this distinctness doesn’t imply it is true to life.
Different areas and figures can be distinguished within a blurred and mixed ground without having recognisable boundaries. Taking a different metaphor, for vision – and in painting – it is rare to encounter or seek such boundaries around figures when they ‘blend into’ a background. Distinctness does not necessarily add to the accuracy of the representation or understanding of its subject when compared to merging with surroundings.


Even if the unity of a metabolic process depends on self-generated boundaries, it does not follow that all forms of autopoiesis or, depending on the definition of autopoiesis, that any form should depend on boundaries of the kind observed and deduced by cell science at a particular time and according to a particular interpretation. The meaning of boundary is not stable across different processes.
In detail, boundary means different things for a cell (membrane) and a body (skin). It means different things for a mind, for a mind and its unconscious, and for an organisation combining many minds (through walls, rules, rewards and enforcement). In practice, distinct boundaries are made by and for observers and for their purposes; for instance, to illustrate a feature in an experiment using super resolution techniques.
This does not mean that a membrane does not exist until it is observed according to a particular technique. It means that the distinctness of the membrane for an observer is dependent on that technique, while the membrane itself is a set of different processes and features, which might not be well-served by being understood as part of a distinct barrier drawn out, in the following image for example, thanks to 4-colour staining and multi-frame reconstruction of high resolution frames:
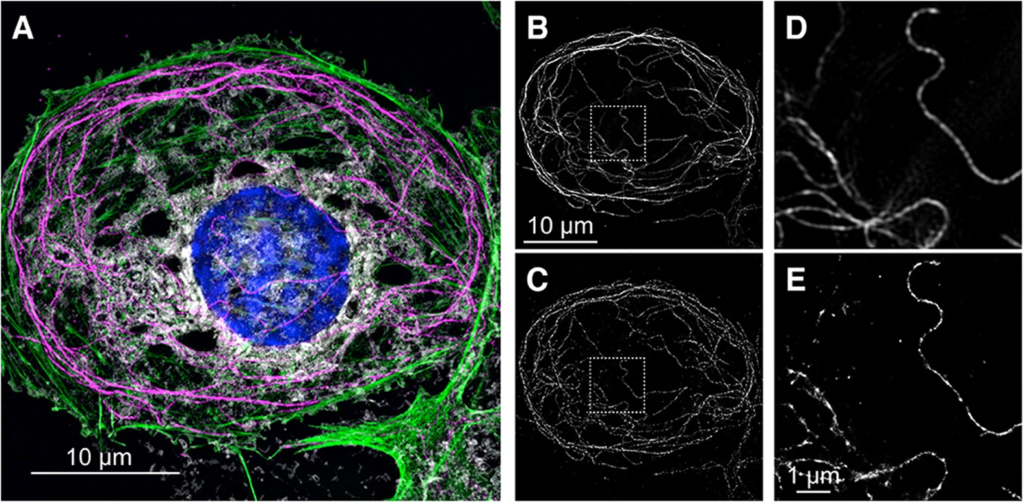
Much more complex than the single cell image of a rat cell, there are many boundaries to a mind and each is well over billions of cells. Which boundary is chosen as the limit for autopoiesis depends on which processes are taken as autopoietic. Will it be the brain sack? The ‘boundary’ of one or many neural networks? The nervous system? The nervous system and all its potential extensions: somatic, technological, social, linguistic, aesthetic and unconscious?
To accurately cover all autopoietic processes, the definition of boundary will have to be much more restricted, or the idea will have to be used analogically – it works like the functioning of a cell boundary – or metaphorically – it’s a boundary of sorts. The first option is rigorous and strict, but also abstract and rational. It depends on logical consistency. The second option is loose and impressionistic, but also prone to imprecision and misrepresentation. It’s a sign and an image, not a faithful correspondence between a concept and a matter of fact.
The same dilemma, between spare consistency and sign-like approximations, holds for other key terms, such as ‘autonomous’. How can cells and cognizers such as language users have the same kind of autonomy? A cell might have closed circuits of production, allowing only energy sources to enter, but a knower or mind uses a language that is itself constantly evolving socially over time. The relative autonomy of a poet, surviving by writing for an audience in a shared language, can only be a weak analogy for the autonomy of the cell.
Thompson constructs his model through empirical expansion, going from a paradigmatic case and applying it more and more widely in order to argue for the importance of autonomy, coupling and intersubjectivity involving unities, their embodiment and enactment. This promotes a general image built on empirically derived autopoiesis:
One of the guiding ideas of this book is that the human mind is embodied in our entire organism and in the world. Our mental lives involve three permanent and intertwined modes of bodily activity – self-regulation, sensorimotor coupling, and intersubjective interaction. Self-regulation is essential to being alive and sentient. It is evident in emotion and feeling, and in conditions such as being awake or asleep, alert of fatigued, hungry or satiated. Sensorimotor coupling with the world is expressed in perception, emotion, and action. Intersubjective interaction is the cognition and affectively charged experience of self and other. The human brain is crucial for these three modes of activity, but it is also reciprocally shaped and structured by them at multiple levels throughout its lifespan.
Evan Thompson (2007) Mind in life: Biology, Phenomenology, and the Sciences of the Mind Cambridge MA: Belknap Press of Harvard University Press, p 243
The embodied mind and Thompson’s idea of autonomous selves enacting their environments leads to a theory of a strong mutual interaction between mind and world. It therefore counters ideas of mind and life as isolated from one another (mind-body dualism) or fully merged with the world (strong materialism). Nevertheless, it leads to other divisions stemming from the concealment of the implications of autopoiesis. This takes place within concepts such as intertwined, coupled, intersubjective and interaction.
For semiology, for a study of the signs at play, there are two contradictory images at work in Thompson’s definition of autopoiesis. One encourages ideas of immersion and reciprocity, no longer a dominance of world by mind, but a more reciprocal togetherness. The other is based on autonomy and independence, of life as self-sufficient, self-produced and self-regulated.
The above passage hides this latter set of signs and implications. Whenever Thompson uses the prefix ‘inter’, it means between an autopoietic process and its environment. It also means between the process and other autopoietic processes. In both cases, autopoiesis is distinguished by a boundary preserving its autonomy. There is an active immersion and yet also a self-producing autonomy:
The first idea is that living beings are autonomous agents that actively generate and maintain themselves, and thereby also enact or bring forth their own cognitive domains. The second idea is that the nervous system is an autonomous dynamic system: It actively generates and maintains its own coherent and meaningful patterns of activity, according to its operation as a circular and re-entrant network of interacting neurons.
Evan Thompson (2007) Mind in life: Biology, Phenomenology, and the Sciences of the Mind Cambridge MA: Belknap Press of Harvard University Press, p 13
The fundamental flaw is that this coexistence of interaction and autonomy is only plausible for the most restricted cases in Thompson’s studies. Once he moves to more and more complex phenomena, the argument becomes analogical and metaphorical. This expansion could still hold were there not contradictions and counter-arguments showing the claims to autonomy to be fragile, given the deep interconnection of processes across autonomy-preserving boundaries.
For loose definitions of autopoiesis, the processes supposed to be autonomous extend into close and distant environments over time. Their past development and future directedness is not within autonomous control, but can be traced back into a shared past, across multiple present dependencies and towards common futures. Entangled processes over time disprove claims to autonomy.