
We can think of autopoiesis as resistance to the imposition of external time. The autonomy of an autopoietic process implies independence from transformational pressures by times outside the process. Autopoiesis should block external time from supplanting the internal time of the process.
It could be objected that everything is resistant to external time, because time is a neutral and arbitrary measure. In that sense everything takes its own time. This can then be placed on a time line of intervals or sequences, but the placing itself doesn’t affect how things happen.
The objection can be dismissed for humans since, for centuries, we have been submitted to an extreme imposition of time on our minds and bodies by various timing systems at work, home and play.
Our experience of external time is not of autonomy but of alienation: ‘This task specifies not only what is to be done but how it is to be done and the exact time for doing it.’ F W Taylor, Principles of Scientific Management, 1911, p. 39.

A response to this dismissal could be that it isn’t a change in time that pressurises us; it’s systems, physical objects, laws, other people, ourselves. This response is not valid if time is a process within things, rather than a neutral external measure. Processes make times.
We are controlled by a timed logic of processes such as internal cycles, growth stages and rhythms: structures of patterns, implications, constraints and implementations. These can be disrupted by, for example, nightshifts, imposed tasks and countdowns. The external intervention is by time processes such as clocks, diaries, alarms, displays and meters, allied to punishments and rewards, such as pay, unemployment, debt, social pressures and advancement.
What goes for humans goes for farm animals – only worse. Farming practices around insemination, gestation, birth, feeding, milking and pastures, around location and length of life, and time of death, impose strict rhythms on animals. Very little autonomy remains for their life processes, independent of the control exercised during their rearing.
Initially and without taking account of definitions of autopoiesis, if we take account of timed processes such as natural and artificial cycles as accelerators and inhibitors of biological processes, the objection about the neutrality of external time can also be rejected for plants; for instance, given the forcing and failure of plant growth according to false seasons.

We can rehearse the debate about humans and time for plants. If time is a neutral and arbitrary measure, then to force plant growth is not to accelerate time, which remains the same. The acceleration is due to differences in light, temperature and nutrients not to a change in time.
The answer is that, for biological lives, time is not an external constant, but rather a multitude of different processes. A change in those processes, such as making them return more frequently, is a cause of the acceleration in – for instance – processes of plant development. For the forced Petunias pictured above, a new time is set by LED lighting. It accelerates their internal growth rhythms.
Excluding potential cosmic events (dust, flares, comet strikes) the most pressing process impinging on internal time for plants is climate change. The accelerations and decelerations in temperature and other seasonal factors have wide effects, such as early budding, flowering and ripening, or droughts, infestations and flooding. The clock determined by temperature change leaves very few plant and animal species untouched.
It doesn’t matter whether some definitions of time give it as arbitrary and abstract. The distinction between internal and external time is about different processes with their own types of time, such as external seasons and internal circadian rhythms. These can exist alongside a common measure of time.
When we suffer from jet-lag, our internal cycles are disrupted by external processes driven by high speed intercontinental flights across multiple time zones. Once we define time as many processes, such as a faster beat or an increasing pressure, we can describe external times as influencing internal ones, while keeping a global definition of time.

A stronger objection could come from physics. If claims for disruption by external time depend on multiple times, then once it can be shown that there is a single objective measure of physical time for all processes, the distinction between internal and external fails and explanations for apparent time disruptions should be about different rates of the same time, not different times and processes.
Measured by radiocarbon dating, objective time can resolve disputes about the age of objects, occurrence of events, length of geological and atmospheric periods, and dates of biological phenomena and features. This doesn’t mean we can’t say something happened faster or slower; for instance, in terms of aging in different epochs as measured according to lifespan.
Carbon dating means there is an objective time whereby periods, speed, acceleration or decay can be measured. It is still possible to make claims about subjective and internal times, but there will be an objective time that takes precedence; for example, in terms of how long something took as opposed to how long it felt.
For the question of external influences on internal time, there is recent research on the possibility of accelerating radioactive decay and hence accelerating time. This doesn’t invalidate objective time. It makes it more complex, since theories of radioactive decay will have to incorporate the idea that ‘… accelerators do not only accelerate particles. For certain nuclei that are unwilling to disappear, they will also accelerate the course of time.’ (p.38)
The external accelerators required mean that this change in the course of internal time will not be commonplace for now. They also point to the role of technology, systems, economies, politics and sciences in different time processes:

If ideas of multiple internal and external times are better explanations and responses to specific problems, such as time-induced crises, objective time as set by physics is no barrier to ideas of internal time. Carbon dating will be able to place events objectively, but phenomena such as enforced acceleration and dictated deadlines are about time-dislocation, not about objective placement.
That something happened without doubt in 1789, tells us very little about the legal and political effects of changing rhythms of life, growth and death for different sections of a population and its environment. Dislocation is a result of multiple processes of time impinging on one another: processes accelerated, decelerated, halted or started by others, with far-reaching effects on the lives caught in them.
The encroachment of external times on internal ones provides explanations for the stress and transformation life forms can be put through during dislocations between time processes. Does the autonomy of autopoiesis support exceptions to, or even refutation of these explanations?
To begin to answer this question and test the robustness of Maturana and Varela’s theory against criticisms based on internal and external time, I’ll now return to their rigorous concept of autopoiesis developed as an explanation of life and as resistance to external effects.
Firstly, since Maturana and Varela define autopoiesis as unity, itself defined as the perfect containment of autonomous processes of self-production, it is not the case that accelerations or decelerations triggered from the outside disprove autopoiesis, if those external changes are not constitutive of that unity.
We might be able to make a process go faster or slower, or even stop it, but this does not mean we have changed the organisation and hence the unity and autonomy of the process. The unity necessary for something to be a particular autonomous machine or life-form remains the same, even when the speed changes.
More broadly, the fact that organisation must be consistent with universal laws – about evolution or chemical reactions or physical interactions – does not discount autopoiesis. This would require a much higher bar, whereby those laws imply that autonomous self-organisation is not possible.
This is not the case, for instance, for accelerators and inhibitors. Neither is it the case for external creation or destruction. The autopoietic thing might come into being or cease to be, it might be accelerated or slowed down, but none of this necessarily changes its defining organisational unity.
Secondly, when taken against Maturana and Varela’s definition of autopoiesis, the critical point that internal processes are subject to external teleology fails. For them, autopoiesis in not defined according to external goals, whether defined by a user or an observer. If external time pressures are taken to have an effect on some target or output decided externally, then this has no pertinence to autopoiesis defined according to logical, structural and dynamic unity.
External time pressures might well change behaviour; inducing distress, for instance. They might well change appearance; making something spin faster to an observer. They might measure effects and products from processes; observing that they have become more or less fruitful over timed increments.
If the structural unity of the process is unchanged, if it retains perfect containment and autonomy as unity, then there is no effect on autopoiesis. A human population might seem beaten into submission under a violent time regime, but it can revolt and reinvent itself rapidly, having retained a deeper unity.

The strength of Maturana and Varela’s definition leads to the following conclusion: It is only when external time processes become necessary for evolving unity that autopoiesis is lost.
This necessity is neither about energy sources, nor about external goals. So long as these do not impact on unity they leave autopoiesis unchanged. Energy might fuel processes of unity; external goals might be imposed on it. The structure of unity might be influenced or even set from outside. So long as unity is autonomous as perfectly contained process, autopoiesis continues.
This resilience appears to be contradictory. How can something be autonomous, if it can be altered from outside? The answer is that it is autonomous and perfectly contained as manner of functioning, like the abstract plan or diagram of a machine. This is how it works, not what it is made of, or what powers it, or its by-product. It might be possible to swap components, inhibit functioning or force outputs. The way of functioning nonetheless remains.
This rationalist and logical version of autopoiesis is resilient so long as unity as the abstract plan is autonomous and perfectly contained. It holds true even if all instanciations of a particular autopoiesis are vulnerable to external change or elimination. However, if there can’t be perfectly contained unity at the abstract level without some kind of external intervention to constitute it, then external time processes can take precedence over autopoiesis.
The manner of autopoiesis is to maintain a topological domain (Maturana and Varela are using this term in a wider sense than its biological use for proteins). An autonomous process of production defines an organisational space: the topology of the process. It is a logical space of functioning that can be mapped.
The domain must not be confused with an environment or habitat. This would negate autopoiesis by diluting its processes into background ones. Instead, the domain is a closed logical space, like the logical space defined for mathematical automata, perhaps known best through the (apparently) simple automata of the ‘game of life‘ as they evolve and progress across a screen.
These domains are representable as images, as is the game of life, but they are more fundamentally defined by algorithms, programmes, theorems and proofs determining autonomous functioning; for instance, in terms of autonomous self-creation and cognition.
Despite connections to automata, in allowing a much wider range of external interactions, Maturana and Varela’s ‘formalism of cellular automata‘ is more complex than those based on mathematical ‘offspring production’, which its authors describe as fragile. This is in stark contrast to the claims for extreme resilience in the biological model:
There exist autopoietic automata which under suitable input conditions could give rise to unbounded complexity growth along the lineages of offspring of such automata (Theorem 4.1). In our setting, this offers a positive answer to the related open problem in the domain of artificial life. On the other hand, Theorem 4.2 shows the fragility of such phenomena — in general one cannot decide whether a lineage will evolve infinitely under given input conditions.
J. Wiedermann / Theoretical Computer Science 383 (2007) 260–269, p 268
Formalism connects the biological to the logical and mathematical; for instance, in an autopoietic process adding to a structure recursively, such as a simple fractal defined by splitting branches but corresponding to plants and ice crystals:
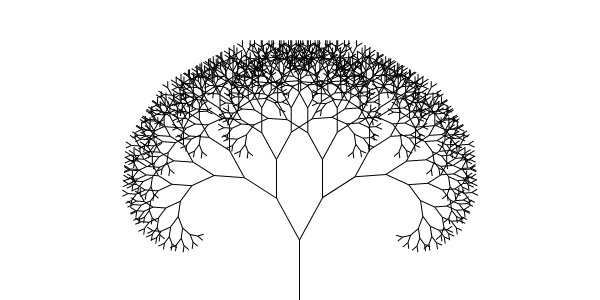

The closure of the domain to external transformational influences is essential for autopoiesis. If the splitting requires external processes as a manner of self-production – if the abstract plan extends externally – then there is no autopoiesis.
When autopoiesis depends on external production, like a host or parasite commanding the type of splitting, or when a computer scientist intervenes – or even can intervene – on a running programme to tweak a fault, then there is no autonomy and no autopoiesis, unless the autopoiesis is extended to include the host-parasite relation, or the scientist.
Extension cancels autopoiesis in both host and parasite. Nonetheless, it can be moved to a higher level including both. The problem with this move is that parasites coexist at ever increasing levels and complexity. In nature, in any empirical case, where is the perfectly contained host-parasite boundary, when it repeatedly shifts and moves ‘at the community scale’?
Parasites that infect new hosts via trophic transmission frequently alter their host’s behavior or morphology in ways that increase predation risk, thereby aiding the parasite in reaching the next host in its life cycle (Poulin et al. 2005). For example, estuarine killifish infected with the trematode Euhaplorchis californiensis exhibit erratic swimming behavior that ultimately makes them up to 30 times more susceptible to bird definitive hosts (Lafferty & Morris 1996). Another trematode endoparasite, Ribeiroia ondatrae, causes amphibians to develop severe limb deformities, including extra or missing limbs (Johnson et al. 1999), which impair the host’s ability to jump and swim, and presumably make them more susceptible to predation by bird definitive hosts (Figure 2). The roles of parasites in predator-prey interactions are rarely obvious, yet they may influence the outcome of trophic interactions at the community scale.
Preston, D. & Johnson, P. (2010) Ecological Consequences of Parasitism. Nature Education Knowledge 3(10):47
Parasitical relations are not troublesome for autopoiesis so long as unity is not affected. This holds even if components or internal relations are changed through external relations. In the above passage, if extension is strictly about trophic relations – exchanging energy (food) sources – then it might be the case that autonomy is maintained at lower levels. If those exchanges involve impairment of unity-denifing processes, then autopoiesis has been breached.
Once again, the bar for impairment is set very high by Maturana and Varela. Their definition allows for substantial changes, such as limbs or relations between them. If the unity of the logical structure remains, then Maturana and Varela’s definition allows for swaps and rearrangements:
The compensatory changes that an autopoietic system may undergo while retaining its identity, may be of two possible kinds according to how its structure is affected by the perturbations: they may be (a) conservative changes in which only the relations between components change; or they may be (b) innovative changes in which the components themselves change.
Autopoiesis and Cognition, p 99
To override autopoiesis, external time processes must impact on the organisational unity of the autopoietic process, in the strong sense of changing it as organisation by breaking that unity; for instance, by extending it through an external process loop necessary for unity.
When a gut depends on the presence of external parasites, autopoiesis as unity of a gut, or unity of an organisation including the gut, is under threat. This is not the case if the parasites merely contribute to energy in the system, or help the gut to function correctly, or even if they are necessary for the gut to work but do not change how the gut functions as unity.
For the necessary breakdown to take place, parasites must challenge how the gut and wider unity are assumed to work according to a perfectly contained plan. That presumed autonomy must be broken and extended to include the parasites such that the manner of functioning of unity is changed.
Even in cases of externally induced change, Maturana and Varela’s concept of ‘coupling’ counters the threat of extended dependencies and takes advantage of them for greater explanatory power:
Whenever the conduct of two or more unities is such that there is a domain in which the conduct of each one is the function of the conduct of the others, it is said that they are coupled in that domain. Coupling arises as a result of the mutual modifications that interacting unities undergo in the course of their interactions without loss of identity. If the identity of the interacting unities is lost in the course of their interactions, a new unity may be generated as a result of it, but no coupling takes place.
Autopoiesis and Cognition, 107
Coupling covers the case of parasitical relations, such as energetic ones, where unity of host and parasite are maintained. Host and parasite can be dependent on one another in coupling, they can even be intertwined, sharing organs and processes. So long as these can be disentangled at the abstract level of their manner of functioning – host this way, parasite that way – then they are each autopoietic even if they cannot live apart and are coupled in a shared domain. However, if their functions are not logically contained, if there is a necessary external reference such as a missing step or an inconsistency requiring an extra element, then autopoiesis fails.
Necessary formal parasitic dependencies can be resistant to empirical evidence giving cause to doubt autopoiesis as resistant to external time. Contingent empirical cases of observed dependence due to intrusion by the times of the parasite or host can provide clues to formal dependence, but they cannot be definitive.
Maturana and Varela’s abstract and rationalist definition always has the option of claiming that any empirical case misses the deeper formal functioning of autopoiesis, such as coupling. Any solution to this impasse turns on classic rationalism versus empiricism arguments about the best explanation of phenomena, with the added complications due to fast evolving fields in biology, AI, technology, sociology, philosophy and cognitive science.
Avoiding dependence on empirical claims, my version of Deleuze’s timed logic sets out speculative metaphysical reasons for the necessity of external relations for any process, thereby rejecting the possibility of any autopoietic resistance to external time. I have defined timed logic here. Given an overview of the timed processes here. With more detail on a timed logic of the present and past, here and here. The explanation of external need applies across all processes. It is a necessity for any timed process.
It could be objected that Maturana and Varela define autopoiesis as a time-independent logical form. This is not correct, since the poetic, self-creating, property of autopoiesis takes place over time. It is an autonomous evolution. Autopoiesis has an internal time of development, in terms of variations over repeated cycles of a given form. The key question is whether that time is formally resistant to external times.
For Deleuze’s timed logic any process determining a present, past and future must extend into all other processes. Any particular present, past or future connects to series of the other times as transformative processes. Furthermore, any process extends into all others such that there are no limits or isolated points or stretches in time. All times connect as continually transformed.
I have highlighted transformation to draw attention to the nature of the extension of processes of time. It isn’t a form of neutral addition, as if components were juxtaposed but left unchanged. It is rather that a process in the present is changed through its relation to the past and to the future. In this timed logic, the whole of the past is concentrated in the present and the whole of the future is anticipated by it.
If we think of autopoiesis as an evolving process, corresponding to a closed yet changing circuit following an autonomous plan, any stage in the process or point on the circuit has a past and a future informing it. Earlier stages prepared for the present one and it prepares for later ones. The challenge of an open timed logic – one that denies any boundary to a timed process – is that each passage between stages opens to the whole of time. Instead of a closed circuit, there is an opening each time there is change or movement along the circuit.
At the most fundamental formal level of the repetition of circuits involving change and continuity, Maturana and Varela’s definition of autopoiesis opens up to external time, because a model of time whereby past, present and future come together through external interventions in order to vary is a better explanation of why there is change in those repeated circuits than one where the repeated circuits change yet remain inviolable.
Each of the core concepts of the most consistent definition of autopoiesis – unity, perfect containment and the closed logical space of a domain – depends on the possibility of unpredictable change, genuine novelty, in a closed system. Yet the best explanation of how this change can take place over time requires unlimited openness and an understanding of how time works through processes that connect past, present and future as determining one another in multiple ways with no pre-set boundaries.